👉 同 步辐 射=XAFS?
👉 同步辐射原位XA FS技术全面解读!
👉 同步辐射软X射线技术解读:原理、应用与案例分析!
👉中科院高能物理所PPT丨同步辐射谱的PCA分析和分峰方法
👉中科院高能物理所PPT丨同步辐射的实验方法
👉四川大学PPT丨同步辐射技术及其在材料学中的应用
👉 “组”传秘方🤣:Artemis进行EXAFS拟合的基本操作
核磁共振( NMR )技术通过观察原子核在磁场中的塞曼分裂现象,可以测量原子核吸收特定频率射频辐射的情况。 由于原子核的共振频率受周围化学环境影响,通过分析共振峰的位置,可以推断出原子核周围的化学键信息,如键长、键角和二面角等,从而帮助构建蛋白质的三维结构 。 NMR 特别适用于研究溶液中的分子结构和动态变化。 NMR 主要适用于小分子量的蛋白质,对于分子量大于 4 万道尔顿的生物大分子, 由于共振峰难以分辨,导致结构解析困难。 此外, NMR 需要将无核磁矩的原子核替换为有磁矩的同位素, 这一过程既昂贵又耗时。 原文链接:
https://doi.org/10.1002/anie.202401343
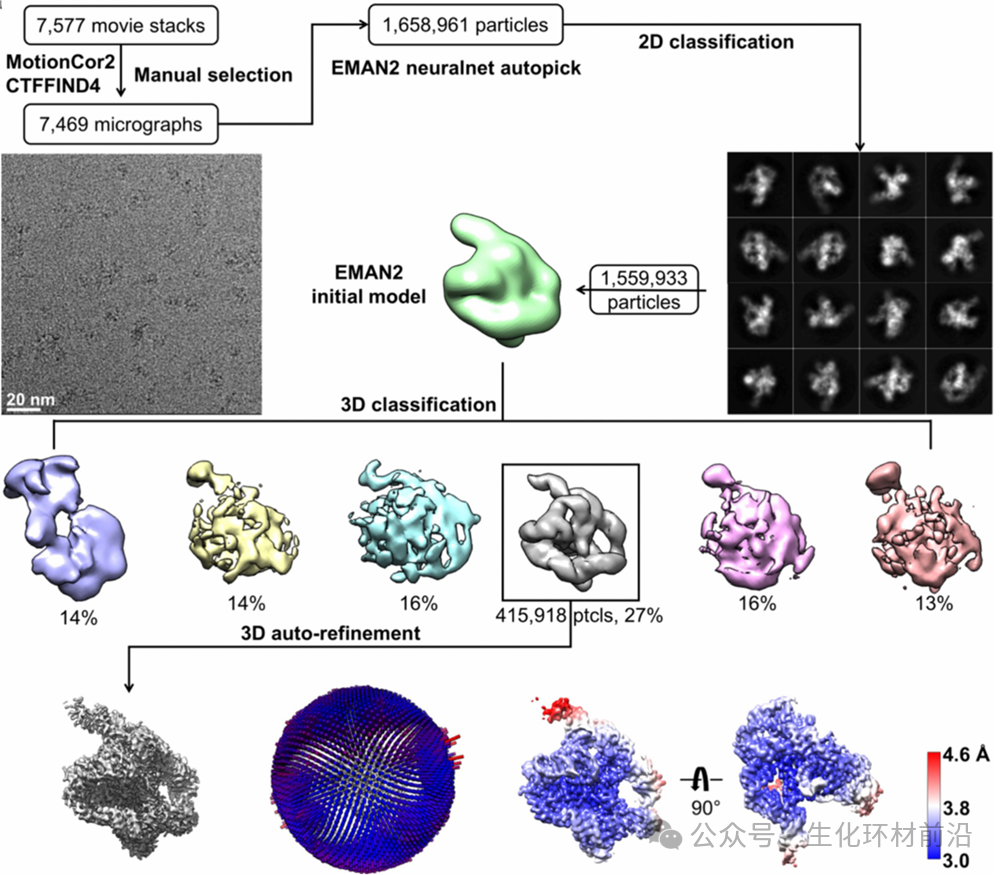
冷冻电子显微学在研究分子量巨大的体系方面具有优势。这种方法可以使用二维晶体,也可以使用单颗粒重建技术来获得三维结构。 然而二维晶体的生长,遇到的困难和生长三维晶体类似,也是一个依靠经验尝试的过程。 利用单颗粒重建技术,需要拍摄很大数量的生物大分子图像,才能够得到足够的信息确定结构的细节。 理论上用单颗粒重建技术得到分辨率为 1 Å 的结构需要处理数以百万计的分子图像,目前还没有完成如此多的图像处理工作的可能性。最近随着新型探测器的发展,在图像质量上得到了大幅度的改善,因此冷冻电镜单颗粒重建获得的结构也接近了原子分辨率, 但是由于电子与蛋白质的相互作用截面很大,多重散射效应使得蛋白质内部结构的分辨率不如表面部分。
原文链接:
https://doi.org/10.1038/s41586-021-03803-w
确定蛋白质分子的三维结构最常用和最准确的手段是 X 射线晶体学。 蛋白质可以在适当的条件下长出晶体,因此衍射技术可以用到蛋白质的结构解析上来。利用晶体学衍射解析结构的历史非常悠久,晶体学早年的工作集中在小分子晶体的结构解析上,一直到二十世纪五六十年代,肯德鲁( Kendrew )和佩鲁茨( Perutz )的工作解析了肌红蛋白和血红蛋白的晶体结构,结构生物学这个今天的 “ 明星 ” 学科诞生了。 目前,利用 X 射线晶体学解析蛋白质结构这门学科也被称为 “ 蛋白质晶体学 ” ,属于 X 射线晶体学的一个分支。之所以在劳厄和布拉格父子的工作完成几十年之后才得到第一个蛋白质的结构, 就是因为蛋白质的结构远远要比小分子复杂,解析蛋白质的晶体结构要困难得多。
原文链接:
https://doi.org/10.1038/s41557-023-01413-9
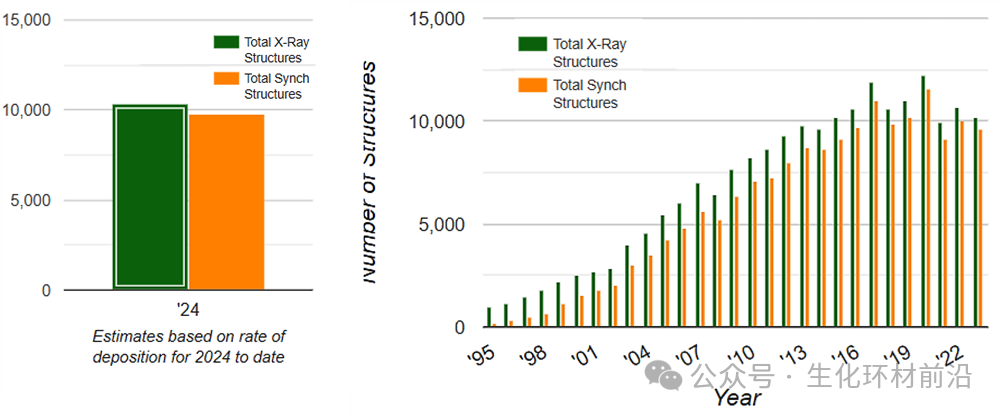
随着 X 射线晶体学的各种理论,以及实验设备的蓬勃发展,现在 X 射线晶体学已经成为确定蛋白质结构的主力技术。到目前为止,在蛋白质数据库( PDB, https://www.rcsb.org/ )中已经收录了超过 10 万个蛋白质结构 ,这些结构中有 9 万多个是用 X 射线晶体学解析出来的,由此可见 X 射线晶体学在确定蛋白质结构中的主导地位。
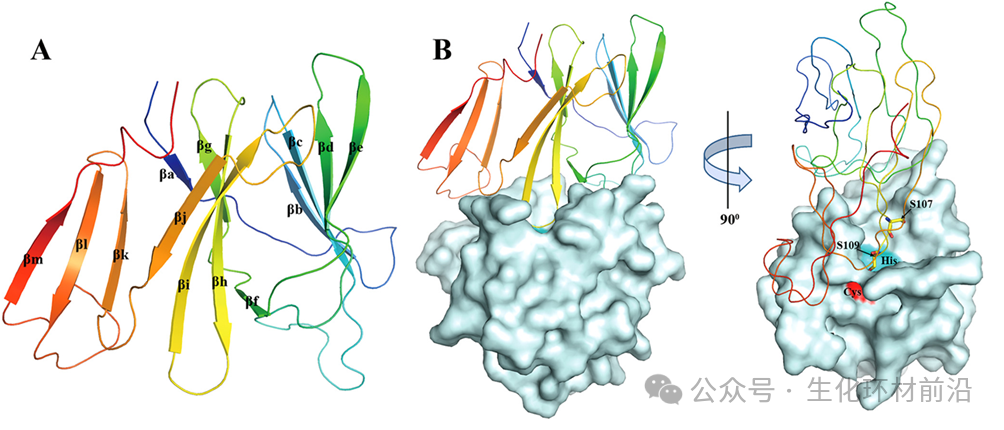
在通过蛋白质晶体学解析的结构中,同步辐射晶体学衍射技术占据了相当大的比例。 根据 Biosync 数据库最新统计( https://biosync.rcsb.org/ ), 2023 年共有 10214 个蛋白质和蛋白质复合物结构通过 X 射线晶体学方法解析,其中 9617 个利用了同步辐射技术。到 2024 年仅半年内,分别上升至 10384 个和 9747 个。 更重要的是, 基于同步辐射结构解析的研究工作已获得 5 项诺贝尔奖 ,分别是 1997 ( ATP 合酶结构), 2003 (离子通道结构), 2006 ( RNA 聚合酶结构), 2009 (核糖体结构)和 2012 年( G 蛋白偶联受体结构)的诺贝尔化学奖。 从数据来看,结构生物学,是目前受同步辐射推动作用最大的学科。 Biosync 统计的利用 X 射线晶体学解析的蛋白质结构数目。绿色为利用 X 射线晶体学解析的蛋白质结构,黄色为利用同步辐射上的 X 射线衍射解析的蛋白质结构。 同步辐射之所以在蛋白质结构中如此重要,是由蛋白质结构解析的特点所决定的, 特别是晶体结构解析中无法回避的相位问题。 相位这一参数无法直接测量,且通过改变晶胞原点位置可以得到不同的结构因子,这些结构因子的振幅相同但相位不同。而相位之间的关系对于确定晶体结构极为重要。
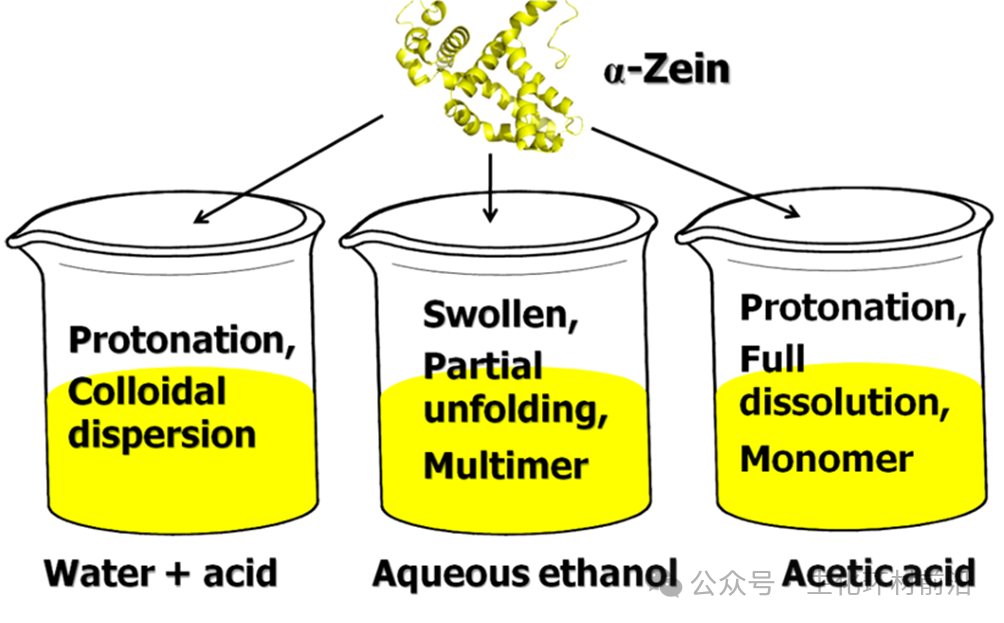
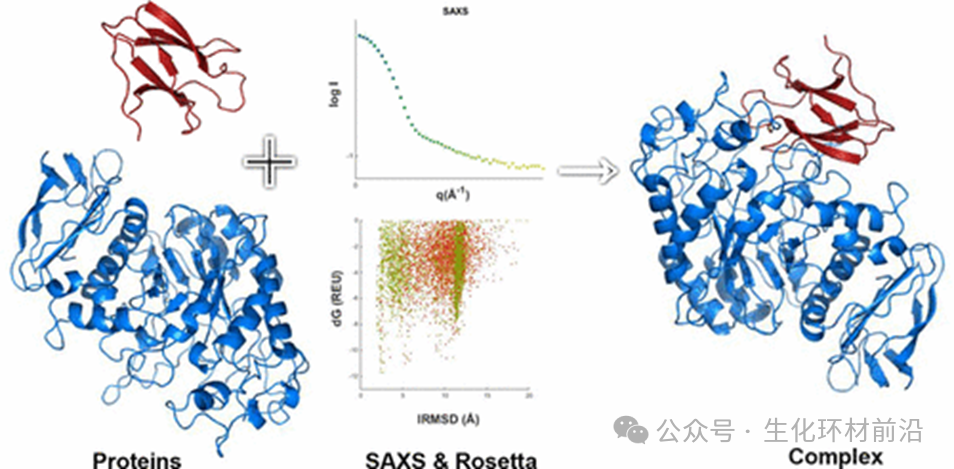
同晶置换法是蛋白质晶体学中用于解决相位问题。该方法通过将重金属离子引入蛋白质晶体而不引起结构变化,获得同晶置换晶体。尽管重原子的加入可能轻微改变蛋白质结构, 但这种方法仍能成功获得结构变化较小的同晶置换晶体。 同晶置换法依赖于制备出同晶置换晶体,这一过程非常困难。 蛋白质结晶本身就困难重重,需要多次尝试,而且重金属离子的引入极易导致蛋白质变性。在同步辐射技术广泛应用之前,解析蛋白质结构是一项艰巨的任务。 直到 80 年代以后,同步辐射作为 SAXS 的光源,提供了极高的入射光强度,有效减小了光斑,实现对样品的结构解析 。 2012 年, Li 等人利用 SAXS 研究玉米醇溶蛋白时,观察到了该蛋白在不同溶剂中的尺寸形状和分散状态,从而得到了蛋白质在溶液中的构象等信息。
原文链接:
https://pubs.acs.org/doi/10.1021/jp305709y
蛋白质复合物结构十分复杂,用 SAXS 解析蛋白质复合物结构时需要联合其他手段。 2013 年 Zhang 等人研究细菌的四型酰 胺酶效应因子和免疫因子形成的复合物时,利用 SAXS 和分析超速离心,观察到该复合物是一个四聚体,同时还发现了这 4 个单体之间相互结合的机制。
原文链接:
https://doi.org/10.1074/jbc.m112.434357
2017 年 Sønderby 等人 研究了 38 对蛋白质复合结构,通过联合 SAXS 和网络对接模拟数据,更加便利 地得到蛋白质复合体在溶液中的构象和三维结构。
原文链接:
https://pubs.acs.org/doi/10.1021/acs.jcim.6b00789
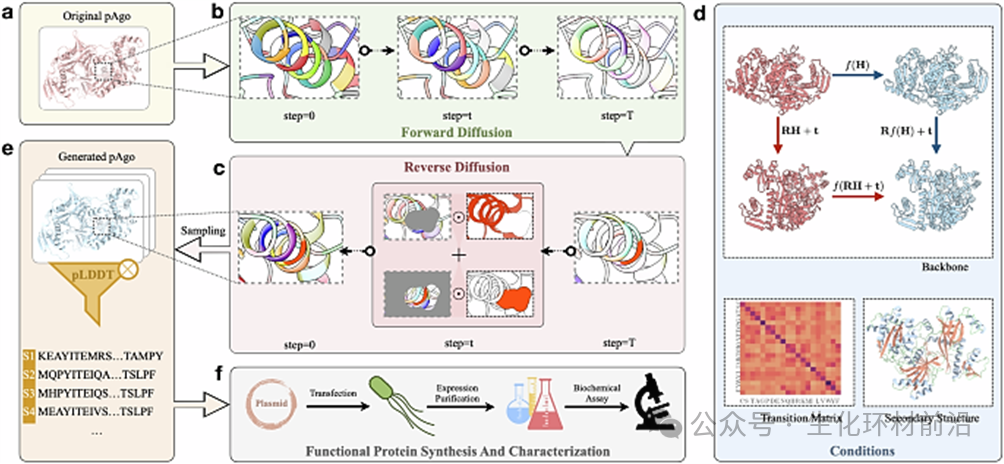
近年来,机器学习领域的热度持续攀升,其影响力已渗透到多个科研领域。 2024 年,上海交通大学自然科学研究院 / 物理与天文学院 / 张江高等研究院 / 药学院洪亮团队等设计了一种扩散概率模型框架,成功设计并生成了具有增强活性的人工程序化内切核酸酶序列。该方法和传统定向进化方法不同,能够一次性改变数百个氨基酸,从而为蛋白质工程选择全新进化起点、生物研究探索蛋白质进化路径、生物科技突破专利封锁等成为可能。 这是迄今为止全球范围内公开文献报道的人工智能生成的结构最复杂,体积最大的单个蛋白,并被湿实验验证有超高活性。 原文链接:
https://doi.org/10.1038/s41421-024-00728-2
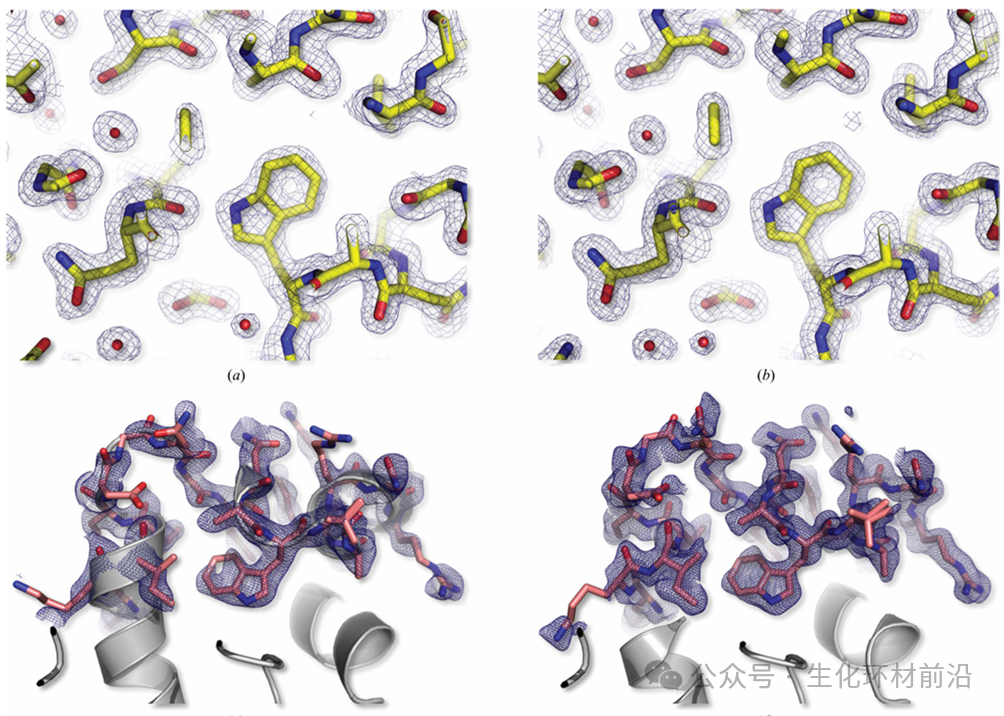
基于同步辐射的串行晶体学 (SSX) 逐渐被重视,且发展迅速。 SSX 用于蛋白质结构解析上具有可在室温下收集衍射数据、能解析微小晶体结构等优点 。 2015 年, Coquelle 等人通过扫描固定域薄硅片上随机取向的晶体,获得母鸡卵白溶解酵素 (HEWL) 微小晶体高分辨率结构信息。在 SSX 的各种研究方法中,最常用的是将晶体固定在微流体芯片等基片上,从而收集基片上每个晶体的衍射数据。
原文链接:
https://doi.org/10.1107/S1399004715004514
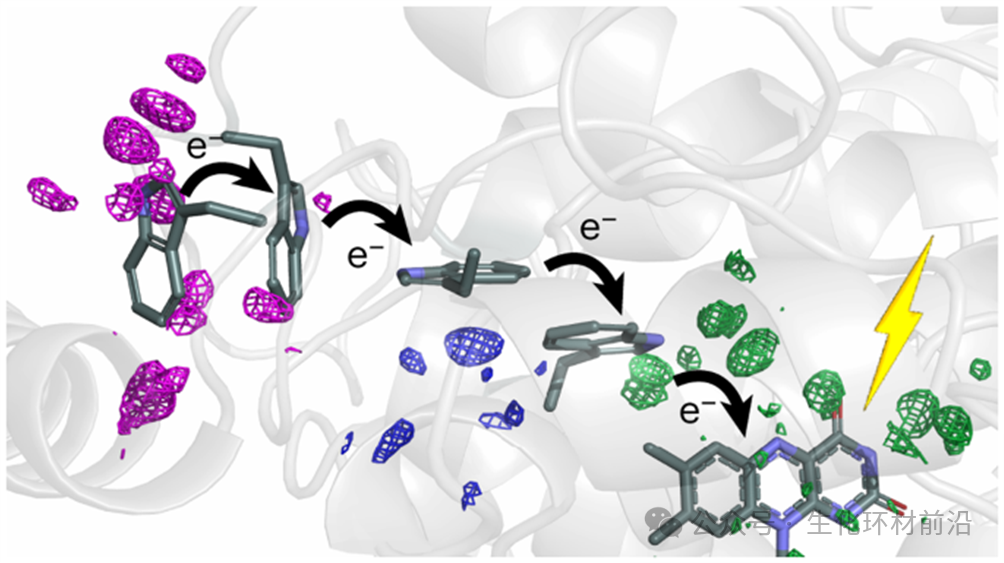
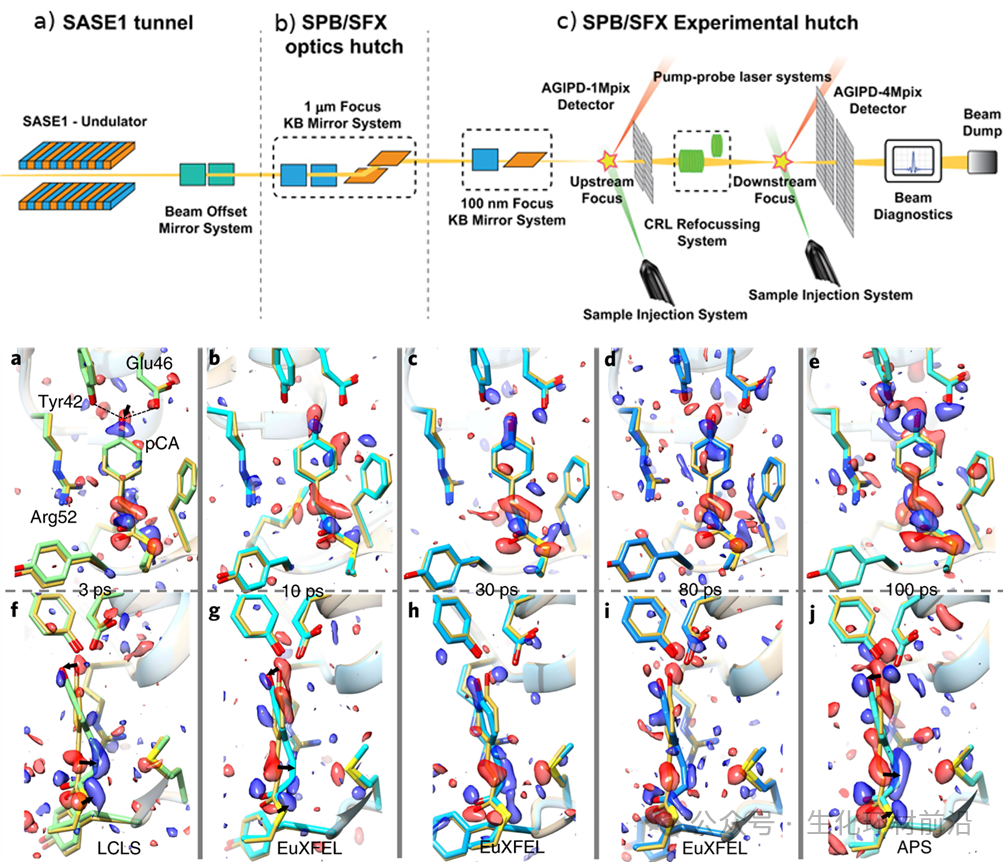
X 射线自由电子激光( XFEL )作为新一代光源,以其极高的峰值亮度和全相干特性,为结构生物学带来了革命性的变化。 XFEL 能够实现“不需要晶体的结构解析”,从而有望克服晶体生长的难题。 2020 年, Grant Mills 在欧洲 X 射线自由电子激光器 上进行了开创性的实验,利用其超亮脉冲和 MHz 脉冲重复率,成功实现了对生物大分子和药物结构的高速和高分辨率确定。
原文链接:
https://doi.org/10.1016/j.ygyno.2015.04.015
随着科学需求的不断增长,新技术如 X 射线自由电子激光和新一代同步辐射光源的出现,未来将对实验装置和实验技术的进一步探究。这些先进的光源技术将为蛋白质结构研究及其他学科的前沿探索带来新的活力和突破。
声明:如需转载请注明出处(华算科技旗下资讯学习网站-学术资讯),并附有原文链接,谢谢!